





Certain plants and nutrients may encourage the production of the enzymes involved in oestrogen metabolism and may support a healthy oestrobolome.
Many of the enzymes required for oestrogen biotransformation are also used in the clearance of xenoestrogens and other toxins. This week we will focus on rosemary, selenium and methylation support.
Rosemary extract
Rosemary is well-established for its anti-microbial, anti-inflammatory and antioxidant properties from the phytochemicals rosmarinic and carnosic acid.
Rosemary induces Nrf2, which signals to the cell nucleus to increase the production of intracellular REDOX capacity and phase II detoxification enzymes. It also increases the expression of genes involved in glutathione synthesis and transport and increases reduced glutathione (GSH) and superoxide dismutase (SOD) levels (1).
In addition, rosemary has been shown to upregulate anti-inflammatory genes, inhibit COX-2 formation, and decrease IL-6 production (2,3). Rosemary reduced the expression of oestrogen and androgen receptors in prostate cancer cell lines and reduced the proliferation of ovarian cancer cell lines (3-6). It also has potent anti-microbial properties, especially to multi-drug-resistant E. coli (7).
Selenium
Selenium is an essential micronutrient, which plays a major role in twenty-five selenoproteins essential for many enzyme systems in the body. These enzymes play a large role in detoxification, anti-inflammatory, and antioxidant-systems (8).
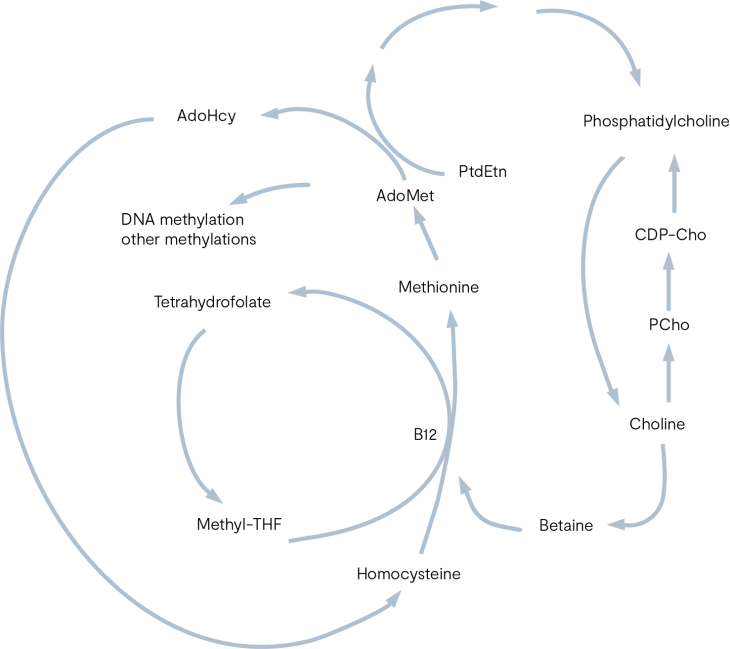
Methylation support: betaine, choline, riboflavin
Some individuals may have epigenetic variants in their enzymes that contribute to inadequate production of methylation co-factors, which may be why some individuals are more prone to the toxic metabolic by-products of oestrogen (10,11).
The enzyme Catechol-O-methyltransferase (COMT) catalyses the methylation of the catechol oestrogens to methoxy oestrogens, which simultaneously lowers the potential for DNA damage and increases the concentration of 2-methoxyestradiol which is an anti-proliferative metabolite (11).
Betaine (trimethylglycine/TMG) is primarily found in seafood, wheat germ and spinach. Inadequate dietary intake of methyl groups can lead to hypomethylation in many important pathways, such as decreasing methionine concentrations and production of S-Adenosyl methionine (SAMe) which is a principle co-factor for many methylation reactions.
Dietary intake of choline can modulate methylation via betaine homocysteine methyltransferase (BHMT), this nutrient (and its metabolite, betaine) regulate the concentrations of S-Adenosylhomocysteine (SAH) and SAMe, which are the main methyl donors for methylation reactions (12,13).